In a past blog post, I have explained in detail the rise of the
diapsids throughout the Late Carboniferous and Early Permian periods.
Unfortunately, we have a rather vague knowledge about what occurred throughout the
rest of the Permian because the fossil record of diapsids is very patchy
between the Kungurian Stage (when Araeoscelis gracilis last lived) and
the latest Permian (when a considerable amount of diapsid fossils reappear at a
few localities across the world).
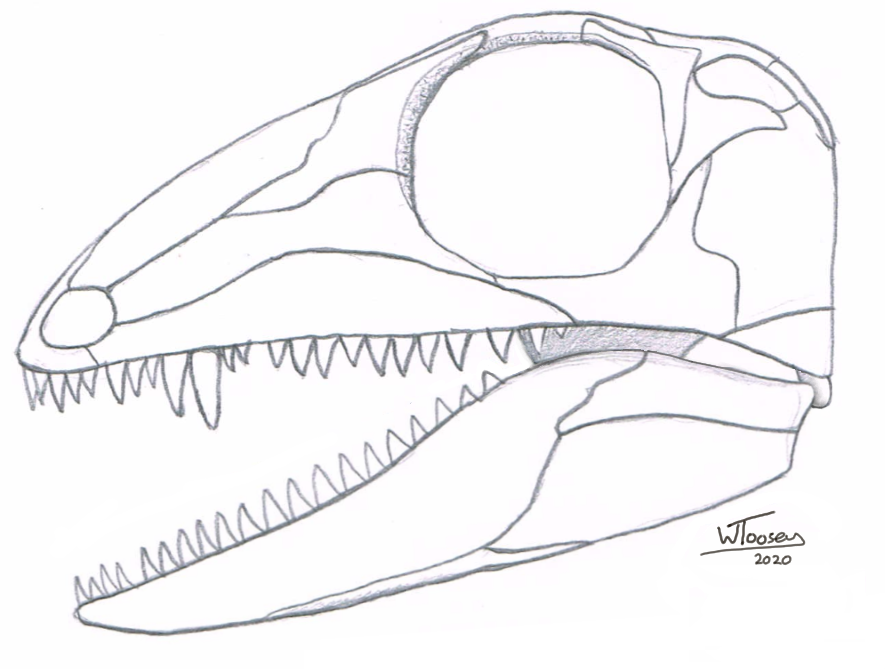
Figure 1: The skull of Lanthanolania ivakhnenko in left lateral
view (drawn from Modesto et al., 2003).

Throughout the whole Guadalupian epoch (Mid Permian), the diapsids
comprise what palaeontologists describe as a “ghost lineage” - this is because
we only know that they were around back then because they are known to have
lived before and after the Mid Permian, although no fossils have ever been
discovered dating to that time. However, not quite: a partial skull found in
the Mezen River basin in the Mezen District of Arkhangel'sk province in Russia
has been described as Lanthanolania ivakhnenko, the genus name which translates
as “forgotten ripper” (Modesto et al., 2003). Of all the hundreds of amniote
fossils found at the Mezen River basin, the only diapsid is Lanthanolania
and was initially misidentified as a specimen of Mesenosaurus, a
synapsid of the varanopid type. Lanthanolania was a miniature neodiapsid,
which may have been no bigger than 30 cm (1 ft) in length as estimated from the
tiny 30 mm long skull.
At the time, diapsids must have been very uncommon, existing in
the relative shadows of much larger anapsids, synapsids and amphibians which
were a lot more successful back during the Permian period. The partial skull of
Lanthanolania has been dated to the ICS Wordian stage, or the uppermost
Kazanian in eastern Europe, somewhere between approximately 266 and 257 million
years ago. Despite the discovery of Lanthanolania inducing a bit of excitement amongst
those experts trying to fill the voids in our knowledge, it still didn’t explain much about
the evolutionary history of diapsids during the Middle Permian because the
remains were so incomplete. One thing we do know for certain is that between
the late Early Permian and the Late Permian, the araeoscelids had already gone
extinct whilst the neodiapsids managed to survive and somewhat diversify.
By the Late Permian, a mixed assortment of neodiapsids
collectively known as eosuchians (meaning ‘early crocodiles’, although this may
be misleading as it didn’t include crocodilians at all) had arrived on the
scene. However, the Eosuchia is something of a wastebasket taxon and the
phylogenetic relationships and taxonomic classification of eosuchians is rather
unclear. It was initially assumed that eosuchians ought to be divided into the
ancestors of archosaurs (e.g. dinosaurs, birds, pterosaurs, crocodilians) and
the ancestors of lepidosaurs (e.g. lizards and snakes), but current thinking is
that their is an even more complex evolutionary history than this. One recently
proposed cladogram (Reisz et al., 2011) demonstrates a supposedly paraphyletic
group with ‘younginiformes’ being the basalmost eosuchians (comprising both
aquatic and terrestrial types), followed by the marine Claudiosaurus germaini
(originally thought of as an early sauropterygian), then by some
representatives of a family of terrestrial lizard-like types known as
‘paliguanids’, and lastly the gliding weigeltisaurids. They all perished by the
Early Triassic, leaving no surviving descendants.
Figure 2: The skull of Youngina capensis in left lateral view
(drawn from Carroll, 1981).

I shall now be covering the diversity of ‘younginiformes’ in
greater detail, commencing with the terrestrial forms. Youngina capensis
(Broom, 1914) was a miniscule lizard-like animal, originally described on the
basis of a fragmentary skull discovered in the Daptocephalus Assemblage
Zone of the Karoo Supergroup in South Africa, dated to the latest Permian at ~
250 million years old. A particular characteristic present in Youngina
is one simple row of osteoderms running midway along the back. Youngina
was part of an ecosystem consisting of a diverse array of fauna such as
synapsids (e.g. biarmosuchians, gorgonopsians, dicynodonts etc.) and anapsids,
in an environment with a semi-arid climate. One amazing find is an assemblage
of five intact, articulated juvenile skeletons, suggesting that Youngina
may have been living in dens (Smith & Evans, 1996). In the subsequent years
following its initial description, a few other specimens of Youngina
were later discovered, predominantly skulls, each being given distinct names
(such as Youngopsis and Youngoides) all of which are now believed
to be synonyms of Youngina.
Figure 3: My reconstruction of Youngina capensis.

The following taxa are also classified as younginids, but with
uncertainty: Heleosuchus
griesbachi, first described as a species of Saurosternon by Richard
Owen in 1876, is known from just one specimen - an incomplete postcranial
skeleton with posterior fragments of the skull. It was first discovered in South Africa from an
obscure horizon, being dated to the Early Triassic or Late Permian. The
specimen was assumed to have been lost until it was moved to the Natural
History Museum of Vienna, Austria (Carrol, 1987).
Galesphyrus capensis (also described by Broom in
1914) is known from an incomplete postcranial skeleton, discovered at the
bottom of the Cistecephalus Assemblage Zone of the Karoo Supergroup in
South Africa, so Youngina is therefore younger.
Kenyasaurus mariakanensis (Harris & Carroll,
1977) is one species of neodiapsid discovered in the Maji-Ya-Chumvi Formation
from Kenya, dated to the Early Triassic. Its phylogenetic relations are
difficult to interpret because only one specimen is known, consisting of very incomplete forelimbs and
pectoral girdle material as well as a missing skull, but may have been closely
related to Lanthanolania. The remains of Kenyasaurus were also
discovered in marine
beds, but lacks adaptations for an aquatic lifestyle, just like Thadeosaurus.
Thus, it was a small, terrestrial lizard-like form measuring roughly 0.5 m long. In addition, Kenyasaurus
was recently found to have not been a member of the younginiformes. With the absence of
well-preserved skull material, knowing the exact phylogenetic relations of all
these taxa has proved to be very challenging.
Figure 4: My reconstruction of Thadeosaurus colcanapi.

Thadeosaurus colcanapi (Carroll, 1981) is a
better-known neodiapsid first discovered in the Lower Sakamena Formation of
southern Madagascar, known from two partial skeletons that are almost complete,
but are lacking the distal (lower) segments of the limbs as well as the skull. Thadeosaurus
was initially and mistakenly presumed to belong to Datheosaurus from
Europe and in turn, Datheosaurus was believed to be a synonym of Haptodus
(a genus of pelycosaur and therefore, a synapsid) but is now considered a basal
caseid within the synapsids. Also bear in mind that two other taxa from the
Permian, Apsisaurus witteri and Heleosaurus scholtzi, previously thought
of as younginiformes, are now believed to have been synapsids of the varanopid
family. It appears that a lot of these primitive diapsids from the Permian were
misidentified as synapsids at first! One other interesting thing about the
genus Thadeosaurus is that it’s merely an anagram of the genus Datheosaurus,
the only anagram of a scientific name in an animal that I currently know of.
Figure 5: My reconstruction of the pelycosaurian synapsid Haptodus baylei.

Thadeosaurus
was a rather small animal, estimated to be approximately 60 cm in length, and
was very lizard-like in appearance, being distinguished by its particularly
elongated tail. Several specimens of Thadeosaurus are known, which
include juveniles (the only known skull is from a juvenile too), most of them
originally being misinterpreted as Tangasaurus mennelli from
northeastern Tanzania (Currie & Carrol, 1984). Thadeosaurus shows no
clear specialisations for swimming despite being discovered in beds deposited
in a marine environment, suggesting that it may have been a terrestrial
neodiapsid living along the coast.
Figure 6: My reconstruction of Tangasaurus mennelli, a neodiapsid that must have lived an aquatic lifestyle as indicated by its long, powerful flattened tail.

One intriguing aspect about the Lower Sakamena Formation is that
the marine deposits consist of an extraordinarily large quantity of diapsid reptiles in comparison to all the other
Late Permian formations elsewhere around the world. The age of the deposits has
been determined by the correlation of vertebrate fossils with those from South
Africa as well as by palynological analysis, being dated from the Capitanian
stage of the late Middle Permian to the Wuchiapingian stage of the early Late
Permian. Among the useful index fossils is the procolophonoid parareptile Barasaurus
besairiei, similar in appearance to Owenetta rubidgei. However, Owenetta
was subsequently found in Early Triassic beds too.
References:
Broom, R. 1914. A new thecodont reptile. Proceedings of the
Zoological Society of London B, 84 (4), 1072–1077.
Carroll, R. L. 1981. Plesiosaur ancestors from the Upper Permian
of Madagascar. Philosophical Transactions of the Royal Society of London B,
293 (1066), 315-383.
Carroll, R. 1987. Heleosuchus: An enigmatic diapsid reptile
from the Late Permian or Early Triassic of southern Africa. Canadian Journal
of Earth Sciences, 24, 664-667.
Currie, P. J. 1982. The osteology and relationships of Tangasaurus
mennelli Haughton (Reptilia, Eosuchia). Annals of the South African
Museum, 86 (8), 247–265.
Currie, P. J., Carroll, R. L. 1984. Ontogenetic changes in the
eosuchian reptile Thadeosaurus. Journal of Vertebrate Paleontology,
4 (1), 68–84.
Harris, J. M., Carroll, R. L. 1977. Kenyasaurus, a new eosuchian
reptile from the Early Triassic of Kenya. Journal of Paleontology, 51
(1), 139-149.
Modesto, S., Reisz, R. R. 2003. An enigmatic new diapsid reptile
from the Upper Permian of Eastern Europe. Journal of Vertebrate Paleontology,
22 (4), 851-855.
Olson, E. C. 1936. Notes on the skull of Youngina capensis
Broom. The Journal of Geology, 44 (4), 523-533.
Reisz, R. R., Scott, D. 2002. Owenetta kitchingorum, sp.
nov., a small parareptile (Procolophonia: Owenettidae) from the Lower Triassic
of South Africa. Journal of Vertebrate Palaeontology, 22 (2),
244-255.
Reisz, R. R., Modesto, S. P., Scott, D. M. 2011. A new Early
Permian reptile and its significance in early diapsid evolution. Proceedings
of the Royal Society B, 278, 3731-3737.
Smith, R., Evans, S. E. 1996. New material of Youngina: evidence of juvenile aggregation in Permian diapsid reptiles. Palaeontology, 39 (2), 289-303.