Amniotes (those tetrapods that lay eggs adapted to land,
equipped with a membrane encasing the embryo called an amnion) are commonly
divided into several groups on the basis of the number of skull openings (or
fenestrae) behind the eye sockets. The most successful of these are the diapsids, which developed two
openings on either side of the skull - one upper supratemporal and one lower
infratemporal fenestra. Practically all living tetrapods that we refer to as
‘reptiles’ have diapsid ancestry: snakes, lizards, crocodilians, turtles, you
name it. They also include the birds (aves), which in turn are descended from
dinosaurs. On the other hand,
mammals along with their reptilian ancestors form the synapsid
clade of amniotes; they have just one opening, which is the lower infratemporal
fenestra. The synapsids and diapsids might have evolved from anapsids (a
primitive clade of amniotes whose skulls lack fenestrae).
Figure 1: Simple pencil diagrams that I did
to demonstrate the various skull configurations of amniotes, all in left
lateral view. (a) diapsid skull of Petrolacosaurus kansensis (drawn from
Reisz, 1981); (b) synapsid skull of Eothyris parkeri (drawn from Reisz
et al., 2009); (c) anapsid skull of Procolophon trigonoceps (drawn from
Romer, 1956). Not drawn to scale.

This differentiation may sound quite straightforward but reflects only the original primitive features and evolution can sometimes cause deception. For example, paleontologists have worked out that plesiosaurs and ichthyosaurs, the most popular Mesozoic marine reptiles, are really highly-evolved diapsids. They are characterised by having only the upper temporal fenestra behind the eye socket, which is known as the euryapsid condition. The euryapsids were initially believed to be a fourth clade of amniotes, but painstaking studies of the known fossil skulls proves that they were really descended from diapsids and that the disappearance of the lower temporal fenestra is just a secondary feature.
One other order of reptiles that
have proven even more difficult to classify are the turtles, which lack
fenestrae behind the eye sockets and were therefore long
regarded as ‘anapsids’. In recent years, though, it now seems thoroughly proven
that they too are heavily restructured diapsids, even though it took
palaeontologists a very long time to work that out. If you look closely at the
skulls of snakes and birds, you’ll find that they are heavily restructured
with both temporal fenestrae lost, but they too are diapsids in accordance with
the basic rules of phylogeny - diapsids are a monophyletic group (consisting of
all descendants of a common ancestor). The diapsids are without doubt the most
successful branch of amniotes with nearly 18,000 living species (when you include
birds), in comparison to the approximately 6,400 species of mammals, which are
the only surviving synapsids.
What exactly did diapsids evolve from
and when? They may have first appeared during the Late Carboniferous from a
clade of anapsid forebearers consisting of taxa like Hylonomus and Paleothyris.
However, it is very difficult to tell for sure due to a patchy fossil record.
The earliest true diapsids are the Araeoscelids, a family of small reptiles
that superficially resembled lizards, the most well-known genera being Araeoscelis
and Petrolacosaurus. The Araeoscelida are known to have lived from the
Late Carboniferous to Early Permian periods.
Figure 2: My reconstruction of Hylonomus lyelli ("Lyell's Forest Mouse"), possibly the world's oldest reptile.

The oldest known diapsid of all is Petrolacosaurus kansensis, whose fossils have been dated to the ICS Gzhelian Stage of the Late Pennsylvanian subdivision of the Carboniferous period (c. 302 Ma). The binomial name translates as "rock lake reptile of Kansas", named after the locality of Rock Lake Shale where the holotype specimen was discovered, which is a partial hind limb. In 1945, it was initially described as a pelycosaur, and therefore a synapsid, but would later be recognised as the oldest known diapsid in 1977 from the discovery and description of new fossils such as a skull showing those two characteristic openings. In a swampy forest environment ruled by giant amphibians, arthropods and insects, Petrolacosaurus was considerably small, possibly measuring around 40 cm (16 in) in length, including the tail.
Figure 3: My reconstruction of Petrolacosaurus kansensis.

Another early diapsid from Kansas that lived around the same time is Spinoaequalis schultzei, discovered in the Calhouns Shale Formation, which also dates to the ICS Gzhelian stage, but slightly younger at ~ 300 Ma. This primitive diapsid is somewhat smaller, at ~ 30 cm (1 ft) and is considered to be an araeoscelid. One fascinating aspect of this animal is the tail, which consists of caudal vertebrae with high haemal and neural spines that are roughly equal-sized (hence the name which means ‘symmetrical spine’), forming a deep, fanned, laterally-compressed tail shape that would be ideal for swimming, which is evident of Spinoaequalis living an aquatic lifestyle. This is additionally backed up by the fact that the fossil was found in freshwater facies amongst the remains of other fully marine animals such as acanthodians (spiny sharks). Yet, the elongated, slender hind limbs are clearly those of a terrestrial animal, so it wasn’t entirely aquatic. Spinoaequalis was therefore among the earliest amniotes to have returned to the water since tetrapods first started to conquer dry land.
Figure 4:
My reconstruction of Spinoaequalis schultzei.

First
described as a lizard in 1910, Araeoscelis sp. dates back to the Early
Permian. Like the contemporary Petrolacosaurus, it too had the same
lizard-like morphology, but had a larger skull with more massive, blunter teeth
specialised for processing insects with tough exoskeletons. Araeoscelis
was also slightly bigger, estimated to have a total length of ~ 60 cm (2 ft),
even though the tail is not known. The lower temporal fenestra is enclosed with
bone which makes the skull more robust, a possible adaptation for anchoring
stronger jaw muscles to help feed on a specialised diet of insects. It
therefore appears to have the ‘euryapsid’ skull configuration, which would later
be so commonplace in large Mesozoic marine reptiles.
Figure
5: The reconstructed skull of Araeoscelis
gracilis in left lateral view (drawn from Reisz et al., 1984).
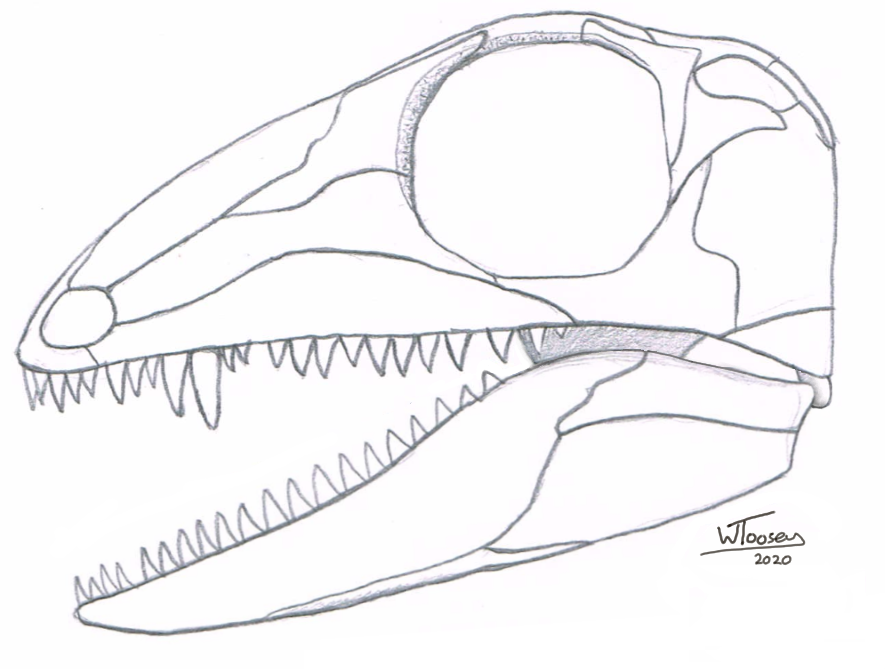
There
are two described species of Araeoscelis, both from Texas: A. casei
of the Admiral Formation (Artinskian stage) and the slightly younger A.
gracilis of the Arroyo Formation (Kungurian stage). Both species are very
much alike
and only appear to be recognised as separate species by differences in age, A.
gracilis being preserved in rocks aged ~ 275 Ma, which is ten million years
younger than A. casei.
Figure 6: My reconstruction of Araeoscelis gracilis.

Other
araeoscelid genera are not well-known, all of them also being dated to the
Early Permian. Zarcasaurus tanyderus, discovered in the Cutler Formation
of New Mexico, has been described from a disarticulated, incomplete skeleton
consisting of a few vertebrae, fractured limb bones and a partial jaw bone
(Brinkman et al. 1984). Another genus closely related to Araeoscelis (as
determined by its considerably long cervical vertebrae) is Dictybolos tener
whose remains have been discovered in the Wellington Formation of Oklahoma (~
290-268 Ma) and may have been a semi-aquatic piscivore; it is estimated to be ~
70 cm long as determined from the isarticulated bones of various
individuals. In Europe, Kadaliosaurus priscus from the locality of
Niederhäslich in Germany is known only from a postcranial skeleton, so its
classification as an araeoscelid is uncertain. Likewise, the partial skeleton of
the somewhat dubious Aphelosaurus lutevensis, known from the Les
Tuilières Formation of Lodève in southern France and first described by Paul
Gervais in 1858, also lacks the skull, ao again it’s difficult to tell if it really
is an araeoscelid.
Figure
7: The skull of Orovenator mayorum in left
lateral view (drawn from Reisz et al., 2011).

There is one other early diapsid distinct from the araeoscelids that dates back to the Early Permian, which is Orovenator mayorum, the most primitive and oldest known neodiapsid (a clade of amniotes consisting of all known diapsids apart from the more primitive types such as araeoscelids). It was first discovered in fissure fills at the locality of Richards Spur in Oklahoma and described on the basis of just two crushed partial skulls and mandibles. It had a rather elongated skull at about 3 cm long, so half the size of Araeoscelis and Petrolacosaurus. The locality of Richards Spur has a characteristic terrestrial vertebrate fauna of about thirty different genera and is presumed to have formed in an upland environment. This is contradictory to the araeoscelids, whose remains have been found in deposits that formed in swampy lowland environments, where fossilisation is more likely. This may mean that the separation of the diapsids into the Neodiapsida and Araeoscelida early in their evolutionary history is a consequence of adapting to life in two contrasting habitats, the neodiapsids in the uplands and the araeoscelids in the lowlands.
Overall, the early diapsids show an astonishing scope of diversity, with some types becoming semi-aquatic, such as Dictybolos and Spinoaequalis, while others like Orovenator were more adapted to drier upland habitats. Still, the fossils of these reptiles are relatively scarce and the fossil record shows a rather large void in them before they reoccur in the Late Permian. The diversity of diapsids would not significantly increase until after the Permian-Triassic mass extinction because throughout the Permian, they lived in the shadows of giant synapsids, anapsids and amphibians. This is very much similar to the small mammals living in the relative shadow of the dinosaurs, for example.
And with that, thank you very much for reading and all the best! Earliest Stages of Diapsid Evolution Part II will focus on the younginiformes...
References:
Brinkman,
D. B., Berman, D. S., Eberth, D. A. 1984. A new araeoscelid reptile, Zarcasaurus
tanyderus from the Cutler Formation (Lower Permian) of north-central New
Mexico. New Mexico Geology, 6 (2), 34-39.
Debraga, M., Reisz, R. R. 1995. A new
diapsid reptile from the uppermost carboniferous (Stephanian) of Kansas. Palaeontology,
38 (1), 199-212.
Lane, H. H. 1945. New
mid-Pennsylvanian reptiles from Kansas. Transactions of the Kansas Academy of
Science (1903-), 47 (3), 381-390.
Olson, E. C., Williams, P. M. 1970.
New and little known genera and species of vertebrates from the Lower Permian
of Oklahoma. Fieldiana: Geology, 18 (3), 359-434.
Reisz, R. R. 1977. Petrolacosaurus, the oldest known diapsid reptile. Science, 196 (4294), 1091-1093.
Reisz, R. R. 1981. A diapsid reptile from the
Pennsylvanian of Kansas. Special Publication of the Museum of Natural
History, University of Kansas, 7, 1-74.
Reisz, R. R., Berman, D. S., Scott,
D. 1984. The anatomy and relationships of the Lower Permian reptile Araeoscelis.
Journal of Vertebrate Paleontology, 4 (1), 57-67.
Reisz, R. R., Godfrey, S. J., Scott,
D. 2009. Eothyris and Oedaleops: do these Early Permian synapsids
from Texas and New Mexico form a clade? Journal of
Vertebrate Paleontology, 29 (1), 39-47.
Reisz, R. R., Modesto, S. P., Scott,
D. M. 2011. A new Early Permian reptile and its significance in early diapsid
evolution. Proceedings of the Royal Society B, 278 (1725),
3731-3737.
Romer, A, S. 1956. Osteology of the
reptiles. University of Chicago Press, Chicago, IL.
Williston, S. W. 1913. The skulls of Araeoscelis and Casea, Permian reptiles. The Journal of Geology, 21 (8), 743–747.
No comments:
Post a Comment