The most popular dinosaur of all time may very well be Tyrannosaurus
rex, but there were many other closely related dinosaurs too, collectively
known as the Tyrannosauroidea, a group of coelurosaurian theropod dinosaurs
consisting of the tyrannosaurids and all their early relatives, being most
closely related to birds. Throughout much of their existence (commencing in the
mid Jurassic some 170 million years ago), tyrannosauroids were mostly small to
medium-sized animals before they attained truly gigantic sizes by the late
Cretaceous. It’s only in the past couple of decades that an impressive number
of early tyrannosauroid fossils have come to light.
The majority of early tyrannosauroids were only under 4 m (13 ft)
long, similar in size to other closely related coelurosaurians. Unlike
the later tyrannosaurids such as T. rex, they all had longer arms and
hands with three fingers as well as lightly built skulls lacking bone crunching
adaptations.
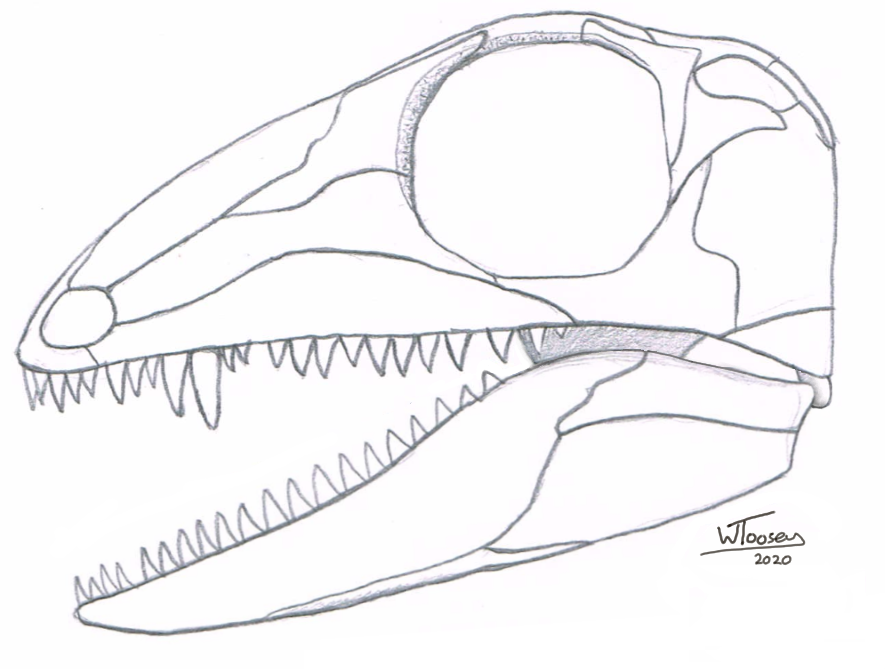
The oldest known tyrannosauroid of all is Proceratosaurus
bradleyi, known from a single, partial skull found in the Great Oolite
Group of the White Limestone Formation at Minchinhampton, Gloucestershire,
England (dated to the Bathonian Stage, Mid Jurassic). From this skull, the
animal is estimated to have been around 3 metres long. What appears to be a
nasal horn on the tip of the skull’s snout was likely just the base of a large,
rounded, mohawk-like nasal crest analogous to Guanlong, to which we now
believe is closely related.
Mistakenly described as a species of Megalosaurus at first by Arthur Smith Woodward in 1910, it was later named as a distinct genus by Friedrich von Huene in 1926, Proceratosaurus meaning “before Ceratosaurus” due to the similar-looking nasal horn-like feature on the skull, akin to the better preserved Ceratosaurus of Late Jurassic North America (von Huene, 1926). Proceratosaurus was assumed to have been ancestral to Ceratosaurus, so both dinosaurs were classified as coelurosaurians. Later in the century, Ceratosaurus was found to have not been a coelurosaur so both dinosaurs weren’t closely related after all, and it wasn’t until 2010 that Proceratosaurus was finally recognised as a definite tyrannosauroid, a member of an evolutionary lineage that would culminate into the T. rex of latest Cretaceous North America (Rauhut et al., 2010).
Figure 1: My reconstruction of Proceratosaurus bradleyi.

One small tyrannosauroid from the mid Jurassic is Guanlong
wucaii, known from two partial skeletons discovered at the bottom of a pit
several feet deep in the Shishougou Formation of far western China, presumably
a sub-adult trampled on by an adult after death. They were both preserved in
sediment formed of volcanic ash and mud, so a natural disaster must have taken
place at the time those individuals died. The name Guanlong means
“crowned dragon” in reference to its thin, fragile, mohawk-like head crest,
which most likely served as a display ornament for intimidating rivals and/or
attracting mates. It was an agile, slender predator, with long, thin legs and a
tail extending beyond the body for balance (Xu et al., 2006).
However, it was not the apex predator of its time and place, which
may have been the same for other primitive tyrannosaurs like Kileskus
from Russia and the dog-sized Dilong paradoxus from China.
Figure 2 My reconstruction of
Kileskus aristotocus (the nasal crest is only a matter of speculation).

Kileskus aristotocus is the oldest known Asian
tyrannosauroid, discovered in rocks dating to the mid Jurassic (Bathonian) from
the Itat Formation of Krasnoyarsk Krai, Russia. It was named and described in
2010 and is known from
a partial skeleton including the front of the snout, a fragment of the lower
jaw, a tooth and some random hand and foot bones. These scrappy remains have
made it difficult to estimate the length of the animal, but may have only been
about 7-8 ft long. It must have lived as a small predator hunting lizards,
salamanders and small mammals in the underbrush, so it would have been rather
puny in comparison to its distant descendants like T.rex.
Even though the fossil lacks a nasal crest, the animal is believed
to be a basal tyrannosauroid within the proceratosauridae (related to Guanlong
and Proceratosaurus) because of certain skull characteristics. These
include a shortened ventral margin of the premaxilla (tip of the snout in the
upper jaw) and enlarged external nares (where the nostrils were situated)
(Averianov et al., 2010).
Thanks to the entombing rocks being so fine-grained, the fossils
of Dilong and Yutyrannus from China have both been preserved with a filamentous
fur-like covering intact, which is so common in coelurosaurians. These
protofeathers were
likely used for insulation or for display purposes and also suggest that the
common ancestors of all tyrannosaurs
must have been feathered too.
Dilong paradoxus is the smallest basal
tyrannosauroid (up to 2 m long), known from a partially articulated skeleton
found along with the scattered bones of several other specimens, discovered in
the lake deposits of the Yixian
Formation in Liaoning Province, northeastern China. The protofeathers of Dilong
would have certainly been used for insulation as it was an animal with a small
body mass to surface area ratio, which means it could have lost internal heat
quite easily. Therefore, it must have been an active predator with a high
metabolic rate, relying on being agile enough to hunt prey such as smaller
reptiles (Xu et al., 2004).
At 9 m (30ft) long, Yuytrannus huali is important in being
the largest known theropod dinosaur yet discovered with feathers preserved, also
from the same formation as Dilong. Yutyrannus was one of the
largest tyrannosaurs alive during the early Cretaceous (dated to 124.6 million
years BCE), but not quite as big as T.rex. It is believed to be a basal
tyrannosauroid, as evident from the foot which lacks the specialised middle toe
that’s useful as a shock absorber while running, as well as for supporting the
weight of the animal. It also had three fingered hands, unlike the stubby two
fingered hands of T. rex and its close relatives. It is therefore
assumed that Yutyrannus was among those primitive tyrannosaurs evolving large body sizes
independently from their later tyrannosaurid cousins (like T.rex), so
they weren’t closely related. Yutyrannus has even been hypothesised to
be a proceratosaurid, those tyrannosauroids with rounded nasal crests (Xu et
al., 2012; Brusatte & Carr, 2016).
Figure 3: Size chart of all the early tyrannosauroids from China I’ve already illustrated. From left to right: Dilong paradoxus, Guanlong wucaii, Xiongguanlong baimoensis, Yutyrannus huali. Height of human figure = approx 1.8 m.

One other Chinese tyrannosauroid is Xiongguanlong bohaiensis,
an intermediate form between the earlier, more primitive tyrannosauroids and
the larger, more advanced tyrannosaurids (such as T.rex) of the Upper
Cretaceous. It was first described in 2009 on the basis of fossils consisting
of a skull with a very elongated muzzle (lacking the lower jaw), a complete set
of cervical and thoracic vertebrae, a right femur (thigh bone) and right ilium (largest
part of the pelvis). From this incomplete fossil material, dated to the early
Cretaceous some 112 million years ago, the animal is estimated to be roughly 4
to 4.5 metres long (Li et al., 2009).
All these discoveries in China help support the hypothesis that
the tyrannosaurs must have had their origins in Asia.
Figure 4: My reconstruction of Juratyrant langhami.

Juratyrant langhami (“Jurassic Tyrant”) is a
tyrannosauroid which, like its earlier relative Proceratosaurus, is also
from the Jurassic of England. It is an important theropod dinosaur find that
was discovered in the Kimmeridge Clay Formation of the Jurassic Coast in southern England, dated to the
Tithonian stage of the Late Jurassic (~ 149 million years ago). It is known from an associated partial
postcranial skeleton of a mature individual, consisting of incomplete leg
bones, a few individual cervical and dorsal vertebrae, a complete sacrum and a complete
pelvic girdle. It was first discovered by avid fossil collector Peter Langham
in 1984 and later described as a species of the North American genus Stokesosaurus
by Benson (2008). However, a study by Brusatte and Benson (2013), found that S.
langhami is in fact a new distinct genus of tyrannosaur more closely
related to Eotyrannus than Stokesosaurus. The body length of Juratyrant
langhami has been estimated at 5 m (16 ft) or so.
Figure 5: My reconstruction of Eotyrannus lengi.

Another
one of those European tyrannosaurs is Eotyrannus lengi, discovered in
plant debris clay beds of the Wealden Group (130-125 Ma) in the Lower
Cretaceous Wessex Formation of the Isle of Wight, southern England. Those plant debris
beds may represent flash floods, when lots of wood got stranded on floodplains.
As the smallest, oldest tyrannosaurs are known from Asia and the largest and
last are mainly from North America, Eotyrannus may help explain that
some tyrannosaurs radiated out westwards from their origins in Asia to Europe, early
in their history, then returned to Asia with more advanced species displacing
earlier ones. Eotyrannus lengi is known from just one partial skeleton preserved
as an irregular block, belonging to a juvenile or subadult. This skeleton
consists of a fragmentary skull, parts of the axial skeleton, pelvic and
pectoral girdle material and a few postcranial limb bones. The fused nasal
bones of the skull tell us that it was indeed a tyrannosauroid, but more
primitive. The individual preserved is estimated to be up to 4 metres long, but
may have grown even bigger, how big exactly we just don’t know (Hutt et al.,
2001).
Figure 6: My reconstruction of a running Qianzhousaurus sinensis individual.

One very interesting example of a later, more advanced tyrannosaur
from China (found in a quarry near the town of Ganzhou, after which the
dinosaur was named) is Qianzhousaurus sinensis, nicknamed “Pinocchio
rex” because of its very elongated skull. Unlike all the other genera described
above, Q. sinensis is a tyrannosaurid, a family of large, advanced
theropod dinosaurs that were truly the top predators of their ecosystems and
evolved from smaller tyrannosauroid ancestors. Q. sinensis is known from
a partial skeleton comprising a complete skull (lacking the teeth), pectoral
material, a left tibia, left femur and several vertebrae (Lü et al., 2014).
These remains show us that Qianzhousaurus, along with the closely
related Alioramus, formed an important, abundant clade of large,
long-snouted carnivorous dinosaurs that lived in Asia during the late
Cretaceous, termed the ‘Alioramini’. Q. sinensis was a tyrannosaur that
lived at the same time T. rex roamed North America, existing until the
K-Pg mass extinction that terminated the dinosaurs’ reign.
In conclusion, the tyrannosaurs were able to become so successful
and roam so far and wide the world over because they existed while the
supercontinent of Pangaea was fragmenting, so they could have dispersed freely between
continents via land bridges to end up evolving in isolation over time. Many of the archaic, primitive tyrannosaurs described and illustrated above were able to
exploit ecological niches as small to medium-sized carnivores in the undergrowth, which is something they were quite proficient at.
References:
Averianov, A. O., Krasnolutskii, S. A., Ivantsov, S. V. 2010. A
new basal coelurosaur (Dinosauria: Theropoda) from the Middle Jurassic of
Siberia. Proceedings of the Zoological Institute. 314 (1),
42–57.
Benson, R. B. J. 2008. New information on Stokesosaurus, a
tyrannosauroid (Dinosauria, Theropoda) from North America and the United
Kingdom. Journal of Vertebrate Paleontology, 28, 732-750.
Brusatte, S. L., Norell, M. A., Carr, T. D., Erickson, G. M.,
Hutchinson, J. R., Balanoff, A. M., Bever, G. S., Choiniere, J. N., Makovicky,
P. J., Xu, X. 2010. Tyrannosaur Paleobiology: New Research on Ancient Exemplar Organisms.
Science, 329 (5998), 1481-1485.
Brusatte, S. L., Benson, R. B. J. 2013. The systematics of Late
Jurassic tyrannosauroids (Dinosauria: Theropoda) from Europe and North America.
Acta Palaeontologica Polonica, 58 (1), 47–54.
Brusatte, S. L., Carr, T. D. 2016. The phylogeny and evolutionary
history of tyrannosauroid dinosaurs. Scientific Reports, 6 (20252):
doi:10.1038/srep20252. Accessed 24/06/20.
Hutt, S., Naish, D., Martill, D. M., Barker, M. J., Newbery, P.
2001. A preliminary account of a new tyrannosauroid theropod from the Wessex
Formation (Early Cretaceous) of southern England. Cretaceous Research, 22,
227-242.
Li, D., Norell, M., Gao, K., Smith, N. D., Makovicky, P. J. 2009.
A longirostrine tyrannosauroid from the Early Cretaceous of China. Proceedings
of the Royal Society B, 277 (1679), 183–190.
Lü, J., Yi, L., Brusatte, S. L., Yang, L., Li, H., Chen, L. 2014.
A new clade of Asian Late Cretaceous long-snouted tyrannosaurids. Nature Communications, 5 (3788):
doi:10.1038/ncomms4788. Accessed: 26/06/20.
Rauhut, O. W. M., Milner, A. C., Moore-Fay, S. 2010. Cranial
osteology and phylogenetic position of the theropod dinosaur Proceratosaurus
bradleyi (Woodward, 1910) from the Middle Jurassic of England. Zoological
Journal of the Linnean Society, 158 (1), 155-195.
von Huene, H. 1926. On several known and unknown reptiles of the
order Saurischia from England and France. Annals and Magazine of Natural
History, 9 (17), 473-489.
Xu, X., Norell, M. A., Kuang, X., Wang, X., Zhao, Q., Jia, C.
2004. Basal tyrannosauroids from China and evidence for protofeathers in
tyrannosauroids, Nature, 431 (7009), 680–684.
Xu, X., Clark, J. M., Forster, C. A., Norell, M. A., Erickson, G.
M., Eberth, D. A., Jia, C., Zhao, Q. 2006. A basal tyrannosauroid dinosaur from
the Late Jurassic of China. Nature, 439 (7077), 715–718.
Xu, X., Wang, K., Zhang, K., Ma, Q., Xing, L., Sullivan, C., Hu, D., Shuqing, C., Wang, S. 2012. A gigantic feathered dinosaur from the Lower Cretaceous of China. Nature, 484 (7392), 92–95.